August 30, 2013 feature
Left brain, right brain: Different patterns of cortical interaction
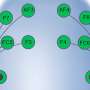
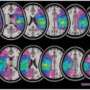
 Homotopic locations in the two hemispheres were identified using relative position on the cortical surface from the centroids of FreeSurfer’s automatically parcellated regions, with 74 regions in each hemisphere delimited by gyral and sulcul boundaries. (Upper Left) The locations of the centroids are shown on the cortical surfaces (small orange circles), and the process of mapping the homotopic location (in blue) of a surface location i in the right hemisphere (in red) is graphically depicted. The homotopic location is the surface node (of 36,002) in the opposite hemisphere with the most similar pattern of distances (geodesic) to the original node (see red and blue traces, Lower Left). (Right) A complete map of homotopic locations for the standard surface model is shown, using color (see key for color map of locations). (B) After identifying homotopic locations, the average within- and between-hemisphere correlation coefficient (ρ) was calculated using resting brain activity at each node and compared at homotopic locations. (Top) Two homotopic seed nodes and their Blood-Oxygenation-Level-Dependent, or BOLD, time series, one in the left hemisphere (LH) and one in the right (RH), along with their corresponding correlation maps in the LH and RH targets (color bar, Right). The correlation maps in the target hemispheres are then averaged over the entire hemisphere (Middle) and stored back at the seed location, separately for the within- and between-hemisphere correlations. (Bottom) The results of this process when repeating for all seed locations, applying Fisher’s z’-transform to yield normally distributed values, and then averaging across all participants on the standard cortical surface (color bar, Right). The first letter in the labels “LL,” “LR,” “RR,” and “RL” indicates a seed location in the left (L) or right (R) hemisphere, and the second letter indicates the target hemisphere. Credit: Copyright © PNAS, doi:10.1073/pnas.1302581110")
(Medical Xpress)—The human brain is divided into two hemispheres – left and right – in which neural functions are said to be lateralized. (For example, language and motor abilities are associated with the left hemisphere, and visuospatial attention with the right.) Although hemispheric lateralization is generally thought to benefit brain function, relationships between lateralization degree and functioning levels have not been quantified. Recently, however, scientists at the National Institutes of Health in Bethesda, MD demonstrated that the two hemispheres have qualitatively different biases: the left prefers to interact with itself – especially for regions associated with language and fine motor coordination – while the right visuospatial and attentional processing regions interact with both hemispheres. Moreover, the researchers provided direct evidence that an individual's degree of lateralization is associated with enhanced cognitive ability.
Dr. Stephen J. Gotts spoke with Medical Xpress about the research that he, Dr. Hang Joon Jo, and Dr. Alex Martin and colleagues conducted – and the challenges they faced in so doing. "One of the tricky things about studying lateralization of function is that it's hard to know exactly which points in the two hemispheres are correspondent." Gotts tells Medical Xpress. This is the case, he explains, because while the hemispheres are roughly symmetrical, there are idiosyncratic differences in cortical folding between left and right for any given individual. In addition, he notes, the exact location of particular folds (known as gyri) varies across individuals.
"Neuroimaging studies have historically adopted a couple of different approaches to deal with this situation," Gotts explains. Some studies, he illustrates, transform the geometry of the brain for each individual into a so-called standard three-dimensional coordinate reference brain – for example, the Talairach-Tournoux atlas. This allows them to estimate symmetrical corresponding points by flipping the left/right x-coordinate about zero. However, he acknowledges that this technique is prone to error by as much as 1-2 centimeters in some brain locations1.
"Another approach," Gotts continues, "has been to compare the magnitude of the neural response in each hemisphere during the performance of a task – for example, a language comprehension task – and calculate a quantitative laterality index to enumerate the extent of lateralization. While this approach makes a lot of sense, and doesn't necessarily require one to solve the correspondence problem, it will be strictly limited to the brain areas that can be activated by the task." In other words, if an area isn't engaged by the task, it's hard to know whether or not it's lateralized. Moreover, it requires many different tasks to be selected in order to address the spatial scope of the entire brain – and Gotts points out that this hasn't been carried out to date.
"Our solution addressed the correspondence problem more directly," Gotts says. The scientists first flattened out a model of each individual's folded cortex onto a smooth surface, spatially warping and stretching each individual brain so that each cortical landmark – that is, gyrus or sulcus – was aligned across individuals. They then found corresponding points in the two hemispheres by their position on this standardized, flattened surface relative to the full set of cortical landmarks. (Sulci are depressions or fissures in the surface of the brain surrounding the gyri.) "Applying the same spatial warping to the functional data then allowed us to compare ongoing, resting brain dynamics between the hemispheres at every position on the cortical surface," Gotts explains.
Utilizing a more traditional, task-based approach to measuring laterality has another downside: researchers typically assess the average magnitude of neural response to a task condition across many individual stimulus events, meaning that dynamical interactions of brain areas aren't as easily assessed. "It's not impossible," notes Gotts, "but to eliminate the effects of stimulus artifacts on connectivity estimates, it requires particular choices of neuroimaging task timing – and it's been done a lot less often than magnitude estimation. The qualitative distinction that we observed in our study between how the hemispheres interact with one another really requires the examination of time-varying neural responses and their co-variation. I don't think that you'd be able to anticipate this finding solely from examining average activity levels."
 and linked to region identity by number (Center and Right) (cluster 1, red; cluster 2, green; cluster 3, light blue; cluster 4, darker blue). Credit: Copyright © PNAS, doi:10.1073/pnas.1302581110")
With respect to the correlations with behavioral ability, Gotts points out that there are probably many different tasks that one could have chosen. "Our choice was to use tasks that have been well-studied and well-normed across individuals as part of the Wechsler intelligence scales – specifically, Vocabulary, which is correlated with many aspects of verbal abilities, and Block Design and Matrix Reasoning, which index aspects of visuospatial processing. These obviously aren't the only possible choices, and it would be nice to follow up this work with a more thorough battery of tasks that would allow us to examine more detailed aspects of language, fine motor control, and visuospatial abilities."
It is important to point out, Gotts adds, that there have been several previous task-based studies that have examined the relationships between lateralization magnitude and cognitive ability, with some reporting a direct relationship as their current study shows. "The main contribution of our study is to demonstrate, at a whole-brain scope, the qualitative differences between the hemispheres in their within- and between-hemisphere interactions. The correlations with behavioral ability really hammer this distinction home, since one needed to use the appropriate metric – that is, segregation versus integration – to see these correlations."
One of the interesting things about the distinction between the hemispheres that the scientists observed, Gotts notes, is that there are implicit hints about it in the literatures on individual cognitive domains. "When people discuss language lateralization, the notion is more like classic modularity: language is operating in the left hemisphere in a manner somewhat isolated, or segregated, from the right hemisphere. This notion may come in large part from the neuropsychological literature, which shows that brain damage to the left hemisphere is much more likely to cause aphasia than damage to the right hemisphere in right-handed individuals.
In contrast, Gotts continues, visuospatial processing and attention involves coordinated processing across the entire visual field, with the left and right halves of visual space represented separately in the right and left occipital cortex, respectively. "Visual processing over the entire visual field requires inter-hemispheric integration, and integration and/or control relates to visuospatial attentional control that is more right-hemisphere lateralized. "Our findings highlight this implicit distinction, making it more explicit and showing that the respective cognitive abilities benefit from it. As a field, I think that we've always assumed that hemispheric lateralization was somehow beneficial for function, but very few brain imaging studies have even examined the issue directly, much less at a whole-brain scope across the range of cognitive domains known to be lateralized."
Moving forward, says Gotts, one of the key outstanding questions is: What is the developmental time course of these hemispheric differences? That is, does the left hemisphere bias for self-interaction exist prior to skilled motor control and language function – or does it emerge later as a consequence of these functions? "If it were to exist prior to handedness and language acquisition in the first few months of age, or even in utero, then the bias could plausibly serve as the cause of the preferential left-lateralization of these functions. One could even try to predict the degree of lateralization present later in life during various tasks, or when at rest, from estimates measured early in life."
A similar set of questions exists for the domain of visuospatial function and the right-hemisphere bias for bilateral interaction, Gotts adds. "Because our method for assessing lateralization only requires measuring resting brain activity and not the performance of complex cognitive tasks, these experiments are actually possible to perform with young infants in a reasonably parallel manner."
According to Gotts, another crucial question for the field of human neuroscience is: What changed from monkeys to apes to humans with respect to lateralization? "Several decades ago, there was the suggestion that monkeys exhibit hand preferences like the ones humans exhibit. After much research, it became clear that monkeys are more symmetrical in their brain control of both motor and visuospatial function. However, apes – such as chimpanzees – appear to be a different story. They appear to exhibit some hand preference lateralization with accompanying brain lateralization, although perhaps not to the extremes to which humans do." (Roughly 80-90% of human males and females are right-handed.) "As with infants, resting brain scans can be performed on monkeys and chimpanzees in a manner similar to those conducted on adult humans."
Regarding other areas of research that might benefit from this study, Gotts thinks it would be possible to apply their methods for assessing lateralization to a range of psychiatric disorders, such as autism and schizophrenia. "There's some suggestion in the literature that lateralization of function is altered in these disorders. Is lateralization qualitatively different from the hemispheric biases we demonstrate for typical individuals – or do they differ in magnitude? We'd also like to understand more about the relationship between handedness and cognitive ability.'
Being left-handed, he illustrates, is associated with a more bilateral representation of language – but this doesn't appear to mandate poorer cognitive abilities in left-handed individuals. "It may be that in left-handed individuals a different optimal weighting or balance of power between the hemispheres is achieved which differs from what we've observed in right-handed males," Gotts concludes. "Our methods could certainly be applied to examine this set of issues."
More information: Two distinct forms of functional lateralization in the human brain, PNAS Published online before print August 19, 2013, doi:10.1073/pnas.1302581110
References
1Quantifying Agreement between Anatomical and Functional Interhemispheric Correspondences in the Resting Brain, PLoS ONE 7(11): e48847 (2012), doi:10.1371/journal.pone.0048847
© 2013 Medical Xpress. All rights reserved.