A canine model of juvenile dermatomyositis
 and scarring on Lorelei’s face. In this photo, taken at 1.5 years of age, lesions on her distal extremities (paws and tail tip) are also evident.")
Human juvenile dermatomyositis (JDM) is the most common childhood inflammatory myopathy (a type of disease affecting muscle and related tissues), causing a characteristic skin rash and progressive muscle weakness beginning around age seven. The causes of the disease are multifactorial, and identifying genetic factors has proven challenging in heterogeneous human populations. The domestic dog suffers from many of the same genetically complex diseases that affect humans, but dogs possess a unique population structure that makes it easier to use genome-wide efforts to tease apart genetic factors. Out of hundreds of genetically isolated dog breeds, only collies and Shetland sheepdogs are routinely affected by dermatomyositis (DMS). This indicates a strong heritable component for this naturally-occurring model of JDM. On the cover of PLOS Genetics' February issue, a four-month-old Shetland sheepdog named Lorelei exhibits facial lesions characteristic of DMS.
In the associated featured article, we first investigated alleles of the dog leukocyte antigen class II genes (located on chromosome 12) that function in immune response, because the human homologues of these genes have alleles associated with JDM. We found very low diversity at three adjacent genes – collies are nearly monomorphic, while Shetland sheepdogs possess the same major haplotype and only one other haplotype worthy of note. Still, we were able to discern that healthy Shetland sheepdogs more often possessed the minor haplotype, rather than the major one. We concluded that the major (predominant) haplotype of both breeds is one risk factor for DMS.
Through genome-wide association analyses, we identified two novel loci on chromosomes 10 and 31 involved in the development of DMS. Genome resequencing of affected dogs uncovered a missense change on chromosome 10 in the gene PAN2, which encodes a key protein involved in mRNA turnover. On chromosome 31, we found an insertion/deletion (an 'indel') in a non-coding exon of the gene MAP3K7CL – a poorly-studied kinase gene. The indel variant removes a predicted binding site for a transcription factor involved in immune response.
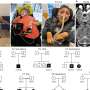
Unexpectedly, the identified allele associated with increased disease risk at each locus is present as a polymorphism in numerous breeds of dog that do not develop DMS. We observed that while affected dogs are homozygous for risk alleles at two or more of these loci, each of these alleles does not individually confer a higher risk of developing DMS. Further, we observed an additive-by-additive epistasis between the PAN2 and MAP3K7CL loci: when one of these loci is homozygous for the risk allele, the addition of a single risk allele at the other locus substantially increases probability of disease.
Lorelei is homozygous for risk alleles at the dog leukocyte antigen genes and PAN2, but is heterozygous at MAP3K7CL. This combination of alleles confers high risk for development of DMS, and 92% of dogs in our study with this genotype were affected. In total, four combinations of alleles are high risk for DMS, five are moderate risk, and the remaining 18 combinations do not confer DMS susceptibility. Only collies and Shetland sheepdogs possess allele frequencies high enough to encourage pairing of risk alleles in combinations that promote DMS.
DMS is also influenced by non-genetic factors. Stressful experiences (e.g., parvovirus infection) commonly precede the first episode of lesions, although many owners of affected dogs cannot identify an overt trigger. In this study, we found that dogs with more risk alleles have a younger median age of disease onset, suggesting that perhaps routine stressors (e.g., rabies vaccination) may suffice for disease development in these cases. Dogs with an intermediate number of risk alleles, like Lorelei, were diagnosed with DMS at a median age of nine months. Lorelei first developed lesions in her ears after a brief illness at two months of age. Upon subsequent puppy vaccinations, lesions and hair loss spread to her eyes and tail tip.
Knowledge of the genetic basis of DMS can now enable selective breeding practices in dogs to prevent future cases and reduce risk allele frequencies. Further, the quality of life for children affected with JDM is markedly diminished by painful, debilitating symptoms, and our work has revealed novel candidate genes and pathways for study. This is the first step toward a deeper understanding of disease pathogenesis, and improved diagnosis and treatment for children.
More information: Jacquelyn M. Evans et al. Beyond the MHC: A canine model of dermatomyositis shows a complex pattern of genetic risk involving novel loci, PLOS Genetics (2017). DOI: 10.1371/journal.pgen.1006604
This story is republished courtesy of PLOS Blogs: blogs.plos.org.