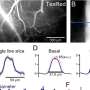
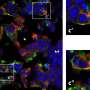
Rodent Somatosensory System. Credit: Neural Coding Lab, School of Psychology, University of New South Wales.
(Medical Xpress)—The constant refrain of the federal BRAIN Initiative can be summarized as the twin-towered call of "tools to codes." To that point, the latest issue of Nature methods is currently running an in-depth series of articles under the title, Focus on Mapping the Brain. The effort seeks to translate the raw numerics of neural structure into the more a satisfying description of "functional connectivity." In the emerging view, the spikes which run on this architecture are not only a language which codes sensory or motor signals, but commands which prescribe how neural circuits grow and change over time. A pioneer in developing tools for optogenetically mapping changes taking place at the synpatic level, is Howard Hughes Medical researcher Karel Svoboda. He was one of the first scientists to definitively reveal the extent of spine plasticity in dendrites. A new paper in Nature Neuroscience, of which Svoboda is corresponding author, seeks to use these new tools to explore the neural coding of somatosensation. To this end, Svoboda and his coworkers have created the highest-fidelity virtual reality system known to date—an illusory mouse whisker sensorium played out on a per spike basis at the millisecond timescale.
In its most primitive form, sensation is often inseparable from the motor action which creates it. The motions of a flexible appendage, such as a cilium or flagellum in a single cell, creates sensations which, in turn, create further motions. It is no surprise that this basic, "poke with a stick," mechanism survives as one of the most sensitive instruments available to creatures as complex as the mouse. Features of the mouse whisker system are known in great detail at the level of cortex. Sensory cells are organized into "barrels" representing each whisker, with up to 2000 layer IV neurons per barrel receiving direct sensory projection. Nearly 80% of these cells respond to most kinds of whisker stimulation, and additionally receive reference copies of the motor commands sent to the whisker by the associated motor cortex.
Using the selective power of Cre recombinase genetic technology, different populations of these layer IV neurons can be targeted by infecting a whole whisker barrel with a modified virus. The virus is then able to express photactivatable channel proteins only those layer IV (L4) neurons that have been targeted. This technique has proven to be very powerful for investigating spike timing in the olfactory system where naturally-delivered odorants can be replaced by precise optogenetic signals higher up in the system. This removes some of the jitter associated with the lower level activation and also permits spike trains to be individually tailored to see their net effect on the output of the olfactory bulb.
This is the exact approach used by Svoboda except that instead of odorant-related signals, whisker-related signals were recreated. By training mice to lick when they encountered a pole in certain positions, the researchers were able to correlate bahavior with the recorded spike trains from cells associated with a particular stimulus. Either stellate cells or inhibitory gaba cells could be selected in different transgenic mice. High speed (1 khz) videography, together with automatic whisker tracing, permitted reconstruction of whisker sweeps, and the associated time of stimulus contact. Not leaving any possible source of timing error to chance, the researcher used a real time version of the linux operating system for control, and even went with ZZ class 0.5 mm gage pins for the stimulus poles. (for those machinists with a neuroscience interest, ZZ class pins are ground to an accuracy of 0.005 mm)
As mouse whiskers sweep at speeds up to 5° per millisecond, such temporal precision in the experimental setup is warranted. Previous studies have shown that L4 neurons can respond to whisker touch in less than 8 ms, with timing jitter on the order of a few ms. Svoboda and his group were able to show that L4 neuron spike latencies could be used to discriminate object locations. When investigated in more detail, their analysis suggested that spike count may in fact be the more robust measure. Temporally precise coupling between spike activity and whisker movement was not required to evoke mouse behavior consistent with the illusory perception of an object in the corresponding target position.
In order for the laser photactivation to convincingly stand in for a real pole, the mouse had to be simultaneously engaged in an active whisker sweep in search of a possible target. Deliberate scrambling of the timing of the activation spikes over the few tens of milliseconds required for a single bout of whisking did not abolish the illusion. This suggested to the researchers that whisker position is unnecessary to discriminate object position. When spike counts were reduced by optogenetically stimulating gabaergic neurons, the mice were biased into indicated no stimulus was detected.
It is likely that the mice were not completely fooled by the artificial stimulus. Other regions of the brain involved in the sensory transmission, like the thalamus, are not directly stimulated by the laser photoactivation. Efforts to assign parts of a stimulus, or subsequent behaviors, to particular features of spike trains is a difficult proposition. As soon as a mouse learns where in the stimulus field to expect a target, its whole world in effect, shrinks to that location. As whisker sweeps are subsequently confined to the target area, spike latency relative to sweep cycle or stimulus, and total spike counts, are likely remapped to new meaning.
Experiments like these push the limits of what is possible with trained animals. The researchers are looking for behavioral results based on changes at the spike level in a limited number of neurons. With trained animals the behavior develops on timescales where spike activity causes significant growth of new and dedicated circuits which support the selected behaviors. There is ample room for many kinds of codes, not just latency and total count, to be supported. As these new techniques are extended to permit a great fraction of the cells in a network to be studied, network level codes will likely emerge to compliment these single cell codes.
More information: Neural coding during active somatosensation revealed using illusory touch, Nature Neuroscience (2013) doi:10.1038/nn.3419
Abstract
Active sensation requires the convergence of external stimuli with representations of body movements. We used mouse behavior, electrophysiology and optogenetics to dissect the temporal interactions among whisker movement, neural activity and sensation of touch. We photostimulated layer 4 activity in single barrels in a closed loop with whisking. Mimicking touch-related neural activity caused illusory perception of an object at a particular location, but scrambling the timing of the spikes over one whisking cycle (tens of milliseconds) did not abolish the illusion, indicating that knowledge of instantaneous whisker position is unnecessary for discriminating object locations. The illusions were induced only during bouts of directed whisking, when mice expected touch, and in the relevant barrel. Reducing activity biased behavior, consistent with a spike count code for object detection at a particular location. Our results show that mice integrate coding of touch with movement over timescales of a whisking bout to produce perception of active touch.
Focus on Mapping the Brain: www.nature.com/nmeth/focus/brainmapping/index.html
Journal information: Nature Methods , Nature Neuroscience
© 2013 Phys.org