May 17, 2013 report
Temporal processing in the olfactory system
The neural machinery underlying our olfactory sense continues to be an enigma for neuroscience. A recent review in Neuron seeks to expand traditional ideas about how neurons in the olfactory bulb might encode information about odorants. One of the main authors, Terry Sejnowski, had the floor for a brief while at last week's national BRAIN Initiative meeting, where discussion of neural codes was a key issue. The Neuron review was published the day after the meeting, and it supports the previously established idea that the olfactory bulb is in many ways structurally comparable to the retina. The authors note however, that due to the apparent sparsity and lack of topographical organization in the olfactory front end, the particular blend of temporal coding used there should differ significantly from that used in the retina.
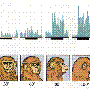
In the quest to link neural activity to function, neuroscientists usually first detect responses to stimuli by looking for an increase in the firing rate of neurons. From these simple rate code beginnings, they usually proceed down a slippery slope involving increasingly contrived measures of response involving the precise timing between the spikes. These temporal codes can be measures like the latency until the first spike, phase offsets, or even "words" of consistently spaced spikes. The prototypical spike word is the recognizable complex spike response found in Purkinjee cells of the cerebellum. More whimsically perhaps, is the idea of coding with grandmother cells, or even more abstract concept cells.
After exhausting such measures, coding at the level of groups of neurons is usually sought in the form or either rich or sparse population codes. Such correlation and synchrony coding links responses across many cells. We recently reported on a theoretical assessment of conditions under which an individual neuron might transition from a rate code to a temporal code. These types of analyses may be particularly relevant in the auditory system where high frequency spiking tracks an auditory stimulus frequency up to several hundred hertz. At some point, when the sound frequency becomes to high for the neuron to phase lock to, the rate code appears to transition into some form of hybrid temporal code.
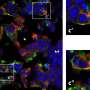
In the olfactory bulb, they neurons which send outputs to the brain are the mitral cells (MT). They would be analogous to the ganglion cells of the retina. Mitral cells receive sensory inputs directly from olfactory receptor neurons (ORNs), which are similar to the photoreceptor cells of the retina. The main difference in the bulb, is that the MTs project directly to the ORNs whereas in the retina, intervening bipolar cells supply the hand-off. It now appears that the tufted cells of the bulb may be analogous to bipolar cells of the retina, and supply significant input to the MTs. We recently discussed the firing patterns of retinal ganglion cells in the recent post on grandmother cells. To understand the differences between the bulb and the retina, we need to look at the firing patterns of the MT cells.
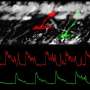
The Neuron article makes the point that counting spikes in the responses of MT cells is often not very helpful. Even when presented with a wide variety of odorants, the average firing rate of MT cells frequently doesn't change—sometimes it even decreases. Also, different odors can result in a similar change in response. Their spontaneous rates are much lower than retinal ganglion cells and may typically be a just few hertz. In studies made in slices of the bulb, the MTs cells appear to have two states, an up state of higher membrane potential, and a low state. The responses of MT cells have also been shown to be phase locked to the respiratory rhythm, and it has been suggested that it is the offset of this phase that encodes the stimulus.
The authors stress that the latency to the first spike has been found to capture a significant portion of the information in a visual stimulus, and similar observations have been made for the bulb. However, studies which claim that particular spike intervals capture a certain amount of information make many assumptions and can be interpreted many ways. Compared to vision, olfactory stimuli are temporally diffusive, and receptors for the same odorant are spread out spatially on the olfactory epithelium. This will cause significant jitter in the responses of ORNs, and in the arrival of inputs to MT cells.
One trick the olfactory bulb has developed is to construct localized glomeruli where individual MT dendrites receive a ball of synapses carrying information about the same odor. How these glomeruli form, and maintain their specificity within a system that continually adds new neurons, is unknown. As it is well established that structure, at least in part, is built and maintained by neural activity, encoding stimuli is not the only thing that spikes do. Since there are many inhibitory projections among MT cells and their glomeruli, researchers have investigated whether they might show some form of the center-surround inhibition that is characteristically seen in the retina. So far none has been found, suggesting other modulatory roles for these projections.
Recent advances in optogenetics permit more discrete components of the olfactory system to be stimulated without the slop involved in using more natural stimuli. In particular, new methods of two-photon infrared excitation can penetrate tissue to excite dye at precise locations. In the visual system the phototransduction pathway is very well understood. By contrast the mechanism used by ORNs to transduce odorants is not. In order to formulate any type of code, it is important to be able to define what it is that needs to be encoded. Applying these techniques will help to unravel some of the ways in which odors, and the incidentals surrounding their detection, represented deeper in the brain.
More information: www.cell.com/neuron/fulltext/S0896-6273(13)00364-4
© 2013 Phys.org