May 10, 2013 report
If you can't beat them, join them: Grandmother cells revisited

(Medical Xpress)—In the absence of any real progress in defining neuronal codes for the brain, the simple idea of the grandmother cell continues to percolate through the scientific and popular literature. Many researchers have reported marked increases in the firing rate of otherwise quiet or idling neurons in response to very specific stimuli, like for example, a picture of grandma. If these experiments are taken at face value, we must accept that grandmother cells, at least in some form, exist. Last December, Asim Roy from Arizona State revived some discussion of this topic with a paper in Frontiers in Cognitive Science. He has just released a follow-up paper in the same journal where he seeks to further extend the idea of the grandmother cell into a more general concept cell principle. A further implication of his paper is that such localist neurons should not be rare in the brain, but rather a commonly found feature.
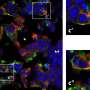
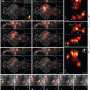
The concept cell derives from an expanding body of research showing that some neurons respond not just to a constellation of stimulus features within a given sensory modality, but also to invariant ideas. For example, researchers have previously reported finding an "Oprah Winfrey" concept cell that could be excited not just by visual percepts of Oprah, but also her name, and even the sound of her name. Roy's new paper suggests that concepts cells would have meaning by themselves, in contrast to neurons in a distributed model, which would represent ideas only as a pattern of activity across a network.
The concept cell theory has been dismissed by many researchers, but represents a valid extremum on the continuity of ways neuron networks can be structured. As such, a theory like this needs to be disproven rather than ignored. Even better then being disproven, a more detailed theory would be welcome. One possible interpretation that reconciles concept cells with distributed network models is to simply have distributed networks of concept cells. When fishing down through the cortex along any given electrode penetration path, it is quite possible to have many quiescent concept cells all around that for whatever reason are not activated at that moment, or are otherwise hidden to the experimenter. Interpreting cells participating in a distributed network as concept cells might just be a lack of sufficient sampling of the relevant network. In that case, the larger reality would be that both viewpoints are just two different interpretations of the same underlying phenomenon.
To get around objections that the idea space is practically infinite while the number of cells that might represent it is finite, Roy notes that concept cells need not be limited to a single concept. At this point, it might be productive to proceed by imagining how concept cells might emerge in a network. For example, would a baby already have grandmother cells? Most would probably argue they don't. A newborn has never seen its grandmother, and although he or she may have some built-in structural hierarchy, that hierarchy has yet to be flashed with very many unique or salient icons. It therefore might be reasonable to assume neurons start out in some kind of distributed mode, but represent little other than perhaps what they experienced in the womb.
When young kids first take up little league baseball or soccer, they generally attempt (at least in the beginning) to maximize their fun such that everyone in the field goes after every ball no matter where it is hit or kicked. Similarly in the newly hatched brain, neurons may quickly learn that spiking at every perturbation that comes its way quickly becomes exhausting. Furthermore, it seems that making synaptic partners indiscriminately must in some way be disadvantageous to the neuron. Competitive mechanisms appear to be in place that link neuron activity and growth to as yet fully defined reward on the molecular level. Such neural Darwinism might simply be the struggle for access to nutrients from the vasculature, like glucose and oxygen, and to dispose of metabolites, like transmitter byproducts. These processes might be enhanced by making the right synaptic partners residing on coveted real estate, and spiking most often at the right time to greatest effect.
As the young athletes learn to adopt more predictive strategies of play, their movements are directed to where the ball is going to be rather than where it is at any given moment. In the extreme, this imperative crystallizes the field into variously named positions with uniquely defined roles and skill sets. Similarly in the brain, the emergence of concept cells could develop over time as a fundamental byproduct of the need to adopt the most energy efficient representations of sensory inputs that map to motor outputs. Included in these sensorimotor hand-offs would be inputs from the body itself, and other expressive or physiologic outputs constrained by the structure of the organism. There are no immediate indications that these transitional representatives in the brain need correspond to real concepts built upon possible activities that can occur in the environment, but there is also no reason why that cannot be the case.
Within the human medial temporal lobe (MTL), up to 40% of the neurons found in some studies have been classified as concept cells. The classification criteria and activity patterns recorded here would warrant closer inspection to draw sweeping conclusions, but some immediate observations have been made. For example, the maximum activation found was reported as a 300-fold increase in spike rate. The background spike rate of a cortical neuron tends to be low, perhaps approaching zero in many cases, so perhaps a better indicator would be an absolute maximum spike rate. We might simply assume a spontaneous background rate of 1 hz for such a cell, and 300 hz for its instantaneous response to an optimal stimulus. We can also ask the following theoretical question: under what conditions does it make sense, from an energetic perspective, for cells within a given network to respond at these relatively fantastic rates to certain rare concepts, while for most others not at all?
Part of the answer may depend on how hard it is for cells to fire at incrementally fast rates, and also how numerous and far away their targets are. Another important consideration is whether the cells can afford to fire at elevated rates on a continued basis without incurring significant damage to themselves. One can even speculate whether there might exist optimal frequencies where possible resonant flow of ions, or overlap of electrical and pressure pulse waves may afford more efficient spiking when high spike rates are called for. In contrast to the cortex, the retinal ganglion cells which comprise the optic nerve tend to fire continuously at relatively high spontaneous rates. Excitatory inputs to retinal ganglion cells result in an increased firing rate while inhibitory inputs result in a depressed rate of firing.
Having a high spontaneous rate gives maximal flexibility and sensitivity for the retina, which is one place where energy expenditure is probably not the major decision point. Another way to look at these cells is that since they can not fire negative spikes, they can effectively double their bandwidth by going with an elevated spontaneous rate in the absence of a stimulus. It is a similar strategy to that often used in electronics for analog-to-digital signal conversion, where bipolar signal sources might not be readily available, and also for small signal amplification in situations where rail-to-tail power sources may otherwise be inconvenient.
In reality, retinal ganglion cell spontaneous rate would probably not be fully one-half that of their maximal rate, but considerably less. A key point to realize is that an important feature of an adaptive system like this is the built-in ability to adjust spontaneous rate across the network according to attention, arousal, and stimulus conditions. This optimizes sensitivity under the dual constraints of the energy available, and the need to eliminate toxic byproducts of using that energy. Whether a neuron can run itself to death by exhaustion, like a racehorse might occasionally do, or whether natural feedback mechanisms in the normal condition would generally prevent this, is unknown. At some point in going inward from the sensory level to the higher cortical areas of the brain, information flow (at least from the retina) transitions to a sparser, lower spontaneous rate environment. At what level, or time, concept cells might begin to appear is only beginning to be unraveled.
Much of the brain can be viewed hierarchically, but there is almost always significant feedback at, across, and among levels. In proceeding hierarchically from sensory to association areas, there seems to be significant convergence from temporal lobe association areas to the hippocampus. The output of the hippocampus then converges, along with other significant pathways from the brain and brainstem, on to particular regions of the interconnected hypothalamus. Ultimately this convergence culminates at specific cells in certain nuclei that convert the electrical currency of the brain into dollops of potent chemical secretions which are active at nanomolar concentrations in the blood.
In the extreme, we could imagine the ultimate concept cells as those few kingpins in certain hypothalamic nuclei controlling things like growth hormone or sex steroid release. These electoral cells spritz appropriately according to both their many far-flung advisors, and to local consensus to control the time and magnitude of each release. Similarly in the deep layers of the motor cortex, the large Betz cells appear to make disproportionately large contributions to motor command to the spinal cord.
Finding these variously incarnated kingpin cells is a major goal in building successful brain-computer interfaces (BCIs), particularly when the number of electrodes is limited. Generally, one does not want to risk stimulating these to death or approaching them too close when trying to hear what they might say. Increasingly, in human experiments, the methods section of the eventual published paper includes statements like, "the subject was then told to focus their thoughts on the target (particular movement)." While no doubt that is a very powerful experimental technique, at this point in time at least, it is also quite vague. Fleshing out exactly what happens when we "focus one's thoughts," is perhaps one the most important research questions of our day.
More information: Paper reference: www.frontiersin.org/Cognitive_ … psyg.2013.00300/full
© 2013 Medical Xpress