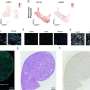
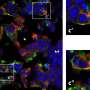
by microarray. Cell lysates were collected for analysis 7 days after treatment. CTRL control. H2O2 hydrogen peroxide. BLEO bleomycin. RAD radiation. Red highlighted, EREG. Agilent microarray data adapted from Sun et al. with permission from Nature Medicine, copyright 2012, Springer Nature. b Representative immunofluorescence staining images (γH2AX and p-53BP1 co-staining, left) and comparative statistics (right) of DNA damage response (DDR) in PSC27 cells treated by DOX (doxorubicin), MIT (mitoxantrone), BLEO (bleomycin), DTX (docetaxel), PTX (paclitaxel) and VBL (vinblastine). DDA DNA-damaging agents (DDAs). NDDA non-DNA-damaging agents. DDR were classified into four sub-categories including 0 foci, 1–3 foci, 4–10 foci and >10 foci per cell. Scale bars, 15 μm. c SA-β-Gal staining of PSC27 cells treated by various agents used in b. Cells were stained 7 days after in vitro treatments. Scale bars, 30 μm. Right, comparative statistics. d BrdU staining of stromal cells treated by different agents as indicated in b and c. Scale bars, 15 μm. Right, comparative statistics. e Quantitative RT-PCR of EREG expression after treatment of PSC27 cells by various agents. Cell lysates were collected for measurement 7 days after treatment. Signals normalized to CTRL. f Immunoblot analysis of EREG expression in stromal cells 7 days after treatments performed as indicated. IC intracellular samples. CM conditioned media. GAPDH, loading control. g Time course expression assessment of a subset of EREG and other typical SASP factors (CXCL8, CSF2, WNT16B, IL6 and MMP3) after drug treatment of stromal cells in vitro. Numeric numbers indicate the individual days after treatment. h Immunoblot measurement of EREG expression at the protein level in the time course described in g. i Comparative appraisal of EREG transcript expression in stromal cells (PSC27) versus cancer epithelial cells (PC3, DU145, LNCaP and M12). Signals normalized to untreated sample per cell line. j Immunoblot assessment of EREG expression in protein lysates of stromal and epithelial cells after bleomycin treatment as performed in i. Data are representative of three independent experiments. ^p > 0.05, *p < 0.05, **p < 0.01, ***p < 0.001. p values were calculated by Student’s t test (c–e, g) and two-way ANOVA (b, i). ^p > 0.05, *p < 0.05, **p < 0.01, ***p < 0.001. Credit: Oncogene (2022). DOI: 10.1038/s41388-022-02476-7")
Genotoxicity induces expression of EREG and other secreted factors of the SASP spectrum in human stromal cells. a Transcriptome-wide profiling of gene expression changes in primary normal human prostate stromal cell line (PSC27) by microarray. Cell lysates were collected for analysis 7 days after treatment. CTRL control. H2O2 hydrogen peroxide. BLEO bleomycin. RAD radiation. Red highlighted, EREG. Agilent microarray data adapted from Sun et al. with permission from Nature Medicine, copyright 2012, Springer Nature. b Representative immunofluorescence staining images (γH2AX and p-53BP1 co-staining, left) and comparative statistics (right) of DNA damage response (DDR) in PSC27 cells treated by DOX (doxorubicin), MIT (mitoxantrone), BLEO (bleomycin), DTX (docetaxel), PTX (paclitaxel) and VBL (vinblastine). DDA DNA-damaging agents (DDAs). NDDA non-DNA-damaging agents. DDR were classified into four sub-categories including 0 foci, 1–3 foci, 4–10 foci and >10 foci per cell. Scale bars, 15 μm. c SA-β-Gal staining of PSC27 cells treated by various agents used in b. Cells were stained 7 days after in vitro treatments. Scale bars, 30 μm. Right, comparative statistics. d BrdU staining of stromal cells treated by different agents as indicated in b and c. Scale bars, 15 μm. Right, comparative statistics. e Quantitative RT-PCR of EREG expression after treatment of PSC27 cells by various agents. Cell lysates were collected for measurement 7 days after treatment. Signals normalized to CTRL. f Immunoblot analysis of EREG expression in stromal cells 7 days after treatments performed as indicated. IC intracellular samples. CM conditioned media. GAPDH, loading control. g Time course expression assessment of a subset of EREG and other typical SASP factors (CXCL8, CSF2, WNT16B, IL6 and MMP3) after drug treatment of stromal cells in vitro. Numeric numbers indicate the individual days after treatment. h Immunoblot measurement of EREG expression at the protein level in the time course described in g. i Comparative appraisal of EREG transcript expression in stromal cells (PSC27) versus cancer epithelial cells (PC3, DU145, LNCaP and M12). Signals normalized to untreated sample per cell line. j Immunoblot assessment of EREG expression in protein lysates of stromal and epithelial cells after bleomycin treatment as performed in i. Data are representative of three independent experiments. ^p > 0.05, *p < 0.05, **p < 0.01, ***p < 0.001. p values were calculated by Student’s t test (c–e, g) and two-way ANOVA (b, i). ^p > 0.05, *p < 0.05, **p < 0.01, ***p < 0.001. Credit: Oncogene (2022). DOI: 10.1038/s41388-022-02476-7
A recent study published in Oncogene and led by Dr. Sun Yu's group at Shanghai Institute of Nutrition and Health (SINH) of the Chinese Academy of Sciences revealed the roles and functional mechanisms of senescence-associated secretory factor epiregulin (EREG) in treatment-damaged tumor microenvironments, providing a new therapeutic target and noninvasive biomarker for treatment of human malignancies including prostate cancer.
Cellular senescence is a unique cellular state with many distinct and unique features. Among these cell characteristics, the unique secretory phenotype of senescent cells, namely senescence associated secretory phenotype (SASP), makes it possible to execute a complex signal transfer function on other cells in the microenvironment.
This secretory phenotype has been found to contribute significantly to the progression of diseases in the elderly, including chronic inflammatory processes. Therefore, the study of SASP associated factors and their functions in tumor and other pathological microenvironments has far-reaching guiding significance for the clinical treatment of geriatric diseases.
In this study, the researchers confirmed that EREG expression was significantly upregulated under senescence induced conditions of DNA damage type by senescence induction in primary human stromal cells. The analysis of cancer samples from clinical prostate cancer and breast cancer patients before and after chemotherapy showed that EREG was significantly up-regulated in senescent paracancer stromal cells.
In terms of the mechanism, DNA damage induces nuclear localization of the transcription factor NF-κB in stromal cells and binds to multiple sites in the EREG promoter region, thereby contributing to the upregulation of EREG expression after cell aging. Other aging-related factors were also noted to promote EREG transcription. In the tumor microenvironment, EREG activates several signaling pathways by binding to EGFR receptors on the surface of adjacent cancer cells, and induces the aggravation of malignant phenotypes such as proliferation, migration and invasion of cancer cells and the occurrence of tumor drug resistance.
Using RNA-Seq analysis, the researchers identified a ubiquitin ligase, MARCHF4, that was significantly upregulated in EREG-stimulated cancer cells. MARCHF4 can regulate the down-regulation of E-cadherin expression in cancer cells and inhibit the apoptosis of cancer cells, thus leading to the occurrence of drug resistance.
In a mouse model, the combination of EREG monoclonal antibody and EGFR monoclonal antibody significantly reduced tumor volume and significantly prolonged the progression-free period. It is worth mentioning that EREG expression level in clinical cancer patients can effectively and accurately predict the efficacy of cancer treatment.
This study elucidated the functions and regulatory mechanisms of a novel senescence-associated secretory factor EREG in the tumor microenvironment and revealed its important value in translational medicine.
More information: Changxu Wang et al, Targeting epiregulin in the treatment-damaged tumor microenvironment restrains therapeutic resistance, Oncogene (2022). DOI: 10.1038/s41388-022-02476-7
Journal information: Nature Medicine , Oncogene
Provided by Chinese Academy of Sciences