June 17, 2013 report
Decoding Rett syndrome: New pieces to the puzzle

(Medical Xpress)—Rett Syndrome is a neurological disorder that affects about 1 in 10,000 girls. Back in 1992, University of Edinburgh researcher Adrian Bird discovered that the protein, MeCP2, plays a major role in the disease. The story of MeCP2 is in many ways a microcosm of human genetics. It has become the showcase gene for many complex epi-genetic phenomena including X-linked inactivation, DNA methylation, and genomic imprinting. These gender-specific bargaining chips provide compatibility in an evolutionary system where sex-chromosome provisioning is inherently assymetric. In two new papers, one in Nature and the the other in Nature Neuroscience, Bird and collaborator Michael Greenberg, show how mutations found in Rett Syndrome affect the interaction of MeCP2 with a key regulatory protein known as NCoR.
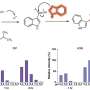
Nearly all cases of Rett Syndrome are caused by mutations at various postions in the MeCP2 gene. Bird and Greenberg analyzed the locations of these mutations using the RettBase MeCp2 database, and found they cluster to two primary locations—the well-known methyl-CpG binding domain, and a new hotspot within a transcriptional repressor domain (TRD). When they compared these locations with mutations found in the general population by using the Exome Variant Server, they found no overlap. This suggests the that the MeCP2 and TRD regions are the primary regions involved in Rett's.
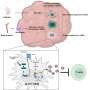
The researchers hypothesized that the newly found TRD region must act through a unknown regulator of MeCP2 function. Using mass spectrometry, they were able to identify several factors which they had purified from Mecp2-EGFP "knock-in" mice. Most of these factors turned out to be subunits of the co-repressor, NCoR, which was previously known to interact with MeCP2. This is the first identified example of a protein-protein interaction known to be disrupted in Rett's.
In the Naturepaper, the researchers further report that activity-dependent phosphorylation of MeCP2 mediates its interaction with NCoR. They used a technique known as phosphotryptic mapping to identify three sites that are directly phosphorylated in MeCP2 as a result of elevation in cAMP or BDNF. More generally, they showed that membrane depolarization, and therefore activity, results in the phosporylation.
One confounding factor in trying to pinpoint the mechanisms underlying Rett Syndrome is that both loss of MeCP2, and overexpression of MeCP2, can lead to the disease. In mouse models of the disease, this could be accounted for by the observation that both loss of NCoR binding, and constitutive binding of NCoR can lead to disease symptoms. While not a complete explanation of the role of MeCP2 in the disease, it provides some clues to help dissect the involvement of the many different kinds of mutations involved.
Despite the rarity of Rett's syndrome, its impact on our understanding of human genetics and neural development should not be underestimated. As one of the autistic spectrum disorders, research on Rett's helps connect molecular mechanics to behavior. For example, when MeCP2 is bound to DNA it can cause condensation of the chromatin structure, and also form complexes with histone deacetylaces. In demostrating that neural activity, and subsequent signal tranduction pathways, lead to modifications of MeCP2, the researchers have revealed a path from the environment directly to the genes.
The X-linked inactivation of one copy of the MeCP2 gene in females adds another layer of complexity to the disease. The celluar mosiac formed by the pattern of inactivation, particularly in the brain, needs more study to be undersatood. The fact that Rett's symptoms can be "rescued" in mice by the expression of MeCP2 in postmitotic neurons is encouraging. In humans, Rett's is frequently not observed untill the first or second year of life. As MeCP2 activation correlates with this period of rapid neural maturation, Rett's is generally considered to be neurodevelopmental disease, as opposed to a neurodegenerative disease.
Rett's is hardly ever observed in males for the simple reason that they fail to thrive long before birth. In those rare cases that a presumably XXY male child is rescued by the additional X chromsome, as in Klinefelder's disease, rare opportunity to study the disease etiology is afforded. The efforts of these researchers, and the larger Rett's community, together with the insights afforded by massive data collation have turned a rare disease into a primary source of knowledge about how evolution proceeds through the interplay of the sexes at the genetic and epigenetic levels.
More information: 1. Rett syndrome mutations abolish the interaction of MeCP2 with the NCoR/SMRT co-repressor, Nature Neuroscience (2013) doi:10.1038/nn.3434 www.nature.com/neuro/journal/v … nt/full/nn.3434.html
Abstract
Rett syndrome (RTT) is a severe neurological disorder that is caused by mutations in the MECP2 gene. Many missense mutations causing RTT are clustered in the DNA-binding domain of MeCP2, suggesting that association with chromatin is critical for its function. We identified a second mutational cluster in a previously uncharacterized region of MeCP2. We found that RTT mutations in this region abolished the interaction between MeCP2 and the NCoR/SMRT co-repressor complexes. Mice bearing a common missense RTT mutation in this domain exhibited severe RTT-like phenotypes. Our data are compatible with the hypothesis that brain dysfunction in RTT is caused by a loss of the MeCP2 'bridge' between the NCoR/SMRT co-repressors and chromatin.
2. Activity-dependent phosphorylation of MECP2 threonine 308 regulates interaction with NcoR, Nature (2013) doi:10.1038/nature12348 www.nature.com/nature/journal/ … ull/nature12348.html
Abstract
Rett's syndrome (RTT) is an X-linked human neurodevelopmental disorder with features of autism and severe neurological dysfunction in females. RTT is caused by mutations in methyl-CpG-binding protein 2 (MECP2), a nuclear protein that in neurons regulates transcription, is expressed at high levels similar to that of histones, and binds to methylated cytosines broadly across the genome. By phosphotryptic mapping, we identify three sites (S86, S274 and T308) of activity-dependent MECP2 phosphorylation. Phosphorylation of these sites is differentially induced by neuronal activity, brain-derived neurotrophic factor, or agents that elevate the intracellular level of 3′, 5′-cyclic AMP (cAMP), indicating that MECP2 may function as an epigenetic regulator of gene expression that integrates diverse signals from the environment. Here we show that the phosphorylation of T308 blocks the interaction of the repressor domain of MECP2 with the nuclear receptor co-repressor (NCoR) complex and suppresses the ability of MECP2 to repress transcription. In knock-in mice bearing the common human RTT missense mutation R306C, neuronal activity fails to induce MECP2 T308 phosphorylation, suggesting that the loss of T308 phosphorylation might contribute to RTT. Consistent with this possibility, the mutation of MECP2 T308A in mice leads to a decrease in the induction of a subset of activity-regulated genes and to RTT-like symptoms. These findings indicate that the activity-dependent phosphorylation of MECP2 at T308 regulates the interaction of MECP2 with the NCoR complex, and that RTT in humans may be due, in part, to the loss of activity-dependent MECP2 T308 phosphorylation and a disruption of the phosphorylation-regulated interaction of MECP2 with the NCoR complex.
© 2013 Medical Xpress