Research identifies protein that signals flowering in squash plants
This research provides some of the most solid evidence to date that FT protein acts as a florigenic signal
The length of the day relative to night, or photoperiod, is a strong determining factor for the induction of flowering in many plant species. Short day (SD) plants require a short day length (or more precisely, a long night) in order to flower. These are plants that flower as the days grow shorter, such as in the fall in temperate regions. Long day (LD) plants will flower when nights are short (and days are long), and typically flower in late spring or early summer. SD crops include rice and maize, and LD crops include wheat, barley, oats and peas. Day-neutral plants will flower under either long or short days. In addition to its fundamental importance in basic plant biology, understanding and manipulating the photoperiodic control of flowering time is an important objective in crop breeding and development programs, because it can aid in optimizing crop yields and other traits for local environmental conditions.
Experimental evidence indicates that a flowering-inducing substance, known as florigen, is produced in the leaves of a plant under inducing conditions, and then is transported through the phloem to the floral meristems, where it acts together with other factors to induce flowering. The precise nature of the florigenic signal has eluded plant biologists for over 70 years, owing to the difficulties attendant with the accurate detection and measurement of compounds in phloem sap, and designing experiments to allow accurate monitoring of long-distance transport of potential signalling compounds in plants. Some of the principal factors associated with the photoperiodic induction of flowering are proteins known as CONSTANS (CO) and FLOWERING LOCUS T (FT). CO does not appear to travel long distances through the phloem, but rather, it strongly influences the production of another substance (i.e. florigen) that does. Recent work in a number of laboratories has focused on a role for FT protein and/or FT mRNA as the florigenic signal, and evidence is building that the FT protein is responsible, at least in tomato, rice, and the model plant Arabidopsis (which is in the mustard or cabbage family). In research published today in The Plant Cell, researchers Ming-Kuem Lin and William J. Lucas from the University of California, Davis, together with a number of coauthors, provide strong evidence that FT protein acts as a florigenic signal in cucurbit plant species (squashes).
The strength of this research lies in the nature of the experimental system utilized. The authors chose Cucurbita squash species because reliable methods for the analysis of translocation through the phloem stream have been developed in this system. However, most cucurbits are day-neutral plants, and to investigate the nature of the florigenic signal effectively, it was essential to utilize an obligate SD plant. In day-neutral plants, it is not possible to manipulate flowering by altering the daylength. By contrast, obligate SD plants will remain in the vegetative state until and unless inducing conditions (such as long nights) occur, and therefore they represent the ideal system for experimental manipulation to investigate this phenomenon. Principal investigator William J. Lucas explained, “it is important for these experiments to use a plant that is 100% inducible; in other words it absolutely will not flower unless it is induced. Then, with carefully designed grafting experiments, we can precisely control the introduction of putative florigenic signals into the translocation stream.” The authors screened nearly one hundred accessions of Cucurbita species to identify one, belonging to the undomesticated squash species Cucurbita moschata, that remained vegetative under LD conditions and flowered only when grown under SD conditions.
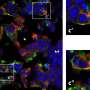
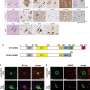
The authors then used the potyvirus Zucchini yellow mosaic virus (ZYMV) as a delivery vector to test whether long-distance movement of FT mRNA and/or FT protein was required for floral induction. The choice of a potyvirus vector was important as these plant viruses do not produce sub-genomic RNA species, whose presence could lead to confounding results. The authors found that C. moschata plants could be induced to flower under the normally non-inducing LD conditions following ectopic expression of FT by the ZYMV vector. Analysis of such induced plants showed that the infection zone of ZYMV was not coincident with the plant’s floral meristems, indicating that FT mRNA is unlikely to be the florigenic signal in this system. Next, heterografting studies were conducted using flowering Cucurbita maxima (pumpkin) stocks (the stock is the portion below the graft union containing the roots) grafted to non-induced, LD-grown C. moschata scions (the portion above the graft union). These experiments led to flowering of the C. moschata scions, indicating efficient transmission of a florigenic signal from the C. maxima stocks through the phloem into the scions. Analyses performed on phloem sap collected from these flowering C. maxima stocks failed to detect the presence of FT mRNA, but revealed the presence of FT protein in the translocation stream.
This research provides some of the most solid evidence to date that FT protein acts as a florigenic signal.
Source: American Society of Plant Biologists