July 17, 2014 report
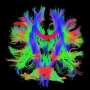
How does the cerebellum work?

(Medical Xpress)—Nothing says "don't mess with me" like a deeply-fissured cortex. Even the sharpest jaws and claws in the animal kingdom are worthless without some serious thought muscle under the hood. But beneath the highly convoluted membrane covering the brains of the evolutionary upper crust hides the original crumpled processor—the cerebellum. How this organ might actually work is the subject of a review published in Frontiers of Systems Neuroscience by researchers at the University of Minnesota.
The Wiki entry under cerebellum does sufficient job of introducing its canonical neural circuit. Centered on the famed Purkinje cell, this basic scheme is present, with telling variation, in the cerebellum of every creature that owns one. As the Minnesota researchers note, the output of all this fuss is an electrically negative signal, "inhibitory" in the neural parlance. To imagine why the cerebellum goes through all this trouble just to apply the breaks we might first look for inspiration as to why electrical circuits in general (on which researchers so frequently model the cerebellum) might use negative potentials.
Whether it is sent with electrons or neurotransmitters every signal has consequences. In the early days of telephony, the standard plug signal lines were called T and R. If you guessed that this stands for transmit and receive, you would be forgiven. The T in this case actually means tip, and the R, the ring. The protruding tip of the plug was always connected to ground for safety in handling while the ring connected to the typical -48 volt lead. But why -48 volts you might ask? As in the cerebellum, the reasons to go negative are many.
One reason negative supplies were the norm had to due with the ease of using germanium PNP transistors in power supplies. For various reasons it a design that uses PNPs is simpler to build when configured as a negative supply. Positive supplies became more common later (as they had been when "valve"circuits were used) when the dominant transistor became the NPN silicium version. Perhaps more importantly, phone companies discovered that with positive voltage on copper wires, there was significant corrosion from electrolytic processes. In the reverse case where the wires carried negative voltage, the copper is "cathodically protected".
As any boat owner knows this is no idle talk. Whenever different metals are placed in conductive liquid you create a battery. When connected together, an electrolytic current will flow which in attempting to equalize the conductivity of the metals removes material from one. Typically in thie case of boats, one adds a piece of zinc to the bottom as a sacrificial anode. Since zinc has a higher effective voltage in the water, electrons will be more inclined to flow from it rather than pitting your propeller.
Your car, (and your zinc-battery-powered flashlight) are usually isolated circuits. Similar corrosive phenomena occur here, and special considerations in grounding the frame (particularly in all-electric vehicles) is warranted. In welding, these same polarity effects are even more acute. If the electrons flow from the workpiece into your small welding tip, it will soon degrade into a blunted mess. If sacrificial welding electrodes are used than the reverse polarity (actually called "straight polarity") can be used, adding the kinetic energy of ballistic electrons to the heat pool of the weld itself.
When any neuron releases transmitter, certain incidentals occur no matter what particular molecule is used to pressurize the released vesicles. Aside from the polarity of the post-synaptic potential that may be elicited, there are all kinds of movements, pulses, and chemical flows that accompany each each fusion event. Purkinje cells are unique in that in one sense they are long-range output projection cells of the cerebellar cortex—akin to pyramidal cells of the cerebral cortex— but in another sense they are like many interneurons in that they use gaba instead of glutamate as their transmitter. But what's in a transmitter you might ask?
Sliding from gaba to glutamate at the synapse is only a short hop in metabolic space. Several enzyme systems can interconvert the two. By comparison, in evolving from insect to vertebrates, the neuromuscular junction at synapses onto muscle has gradually shifted from glutamate to acetylcholine as the transmitter assailant of choice. The enzymatic cycle of synaptic life comes full circle in cells like starburst amacrine cells of the retina which employ both gaba and acetylcholine in signalling. Each transmitter plan includes its own unique chemical baggage and generates, if you will, its own "neural corrosion". Spikes themselves come at price. It's not just the energy they require (and the toxic radicalsoften linked with the mitochondrial source of that energy), but there is also an associated opportunity cost in using one transmitter over another. If instead of excitatory amino acids your cell uses catecholamines, metabolic pathways dependant on tyrosine may often come up short. In other words, in following a Le Chateliers Principle for synapses, many things would need to adjust. The way nervous systems appear to have adjusted is that their neurons, in addition to completing electrical circuits, also complete metabolic circuits.
As we have indicated, perhaps at length, the choice of polarity in electrical power is not a neutral one. Every circumstance will have its own chemical details, and just as important, its own ideal amplitude. When power applications, through optimization, greatly reduce either their current, their voltage, or both, they essentially become signal applications. Here, inverting polarity has more nuanced effect. By way of analogy, when introducing a speaker to a noisy audience, a good moderator sends an inhibitory signal to hush the crowd, effectively making the speaker more clear. The brain appears to do the same exact thing on every scale imaginable.
In the cerebellum, each step in the circuit sends inhibitory projections in both the forward and backward direction in parallel to its own its own signal path. These projections both quiet the arena before the main event, and also turn it off the show at its conclusion. Similar inhibitory local circuits in the retina create the so-called Mexican Hat (if we can still use the term in polite neurophysiology) receptive field found in many of its cells. This field consists a center spot where a stimulus activates the cells, and an inhibitory surround area of activation that acts to quiet it. The net effect is to sharpen the response in space but the same thing can be done in the temporal field as well.
At the level of the synapse, the manipulation of signal polarity has been made into an artform. The channels which carry ion currents have tuned their kinetics so that the largest and sharpest spikes can be made when inhibitory currents precede excitatory currents. This is really just advanced common sense. A side effect of extending the voltage range of channel operation downward, is that a "rebound" spike can be generated even by a purely inhibitory signal acting alone. As the authors note this mechanism is commonly found in cerebellar circuits.
None of this banter can hide the fact that despite a few pockets of eclectic knowledge, not only do we know little about how the cerebellum works, we know next to nothing about what it does. Purkinje cells have a great knack for becoming whatever researchers ask them to become. We know for example, that in a central region of the cerebellum called the floccular complex, Purkinje cells have been found to encode various metrics of the eye during smooth pursuit and other kinds of following movements. Similarly, topographic maps for other parts of the body have been loosley superimposed on the cerebellar surface in a crude intimation of its idealized function. Outputs of the cells in different areas have been imagined to encode everything from limb position, joint angle, speed, direction, or acceleration, to more derived esoterics like muscle force, spindle tension, elongation, fatigue, error, any number of predictors.
The one thing electrophysiologists have kept in their pocket was something called the complex spike. The incidentals of this beast—where it came from, how many spikes it contained, what ion conductances caused it, and so on—was the saving grace that could be turned to in answering all things cerebellar. It now seems, at least according to this new review, than even good old simple spikes can encode many of these variables. What is still going for Purkinje science, however, is the singular nature of the"climbing fiber" that intimately wraps the this cell from head to toe and seems to be critical in controlling thecomplex spike. When this input is activated, it may not just be polarity that is the critical signal variable—the purely physical disturbance of such a large synaptic overload would seem to literally "shake the tree" of the Purkinje.
With such gaps in knowledge now in mind, we might suggest that neuroscience needs to take a closer comparative look at other species to form new hypothesis. There is no better creature perhaps, then electric fish to start the investigation. Electric eels, fish really, can generate lots of electric power to stun their prey. The real masters though, are the Morymrid electric fish that used their powers instead for signaling. These guys, intriguingly, have gigantic cerebellums which presumably handle the needs that arise in electrosensing and communication. Many of their fascinating cellular adaptions include features not found in the cerebellums of mortal animals.
We won't get into them here other than to say that they have not just sprung fully-formed from the void, but rather have been amplified from existing neural wetware to fill existing niches. One thing I think we must ask of such a fish, (and the cerebellum of all species) in light of our knowledge of the thoroughly integrated transport of signal, power, and organelle throughout central and peripheral nervous systems, is what is the cerebellum actually supplying to the muscle? The femoral artery that feeds chemical power to our leg might be 5/8th inch in diameter. Considering that the sciatic nerve which generally is believed to convey only signal is more like 3/4 inch in diameter, it may be delivering more than electrical currency.
More information: The cerebellum for jocks and nerds alike, Front. Syst. Neurosci., 16 June 2014. DOI: 10.3389/fnsys.2014.00113 . http://journal.frontiersin.org/Journal/10.3389/fnsys.2014.00113/abstract
© 2014 Medical Xpress