June 4, 2013 report
Distinguishing REM sleep from other conscious states

(Medical Xpress)—Despite decades of research, little is known about the function of REM sleep, or the dreams that often accompany it. Rapid eye movements occur in most mammals, with a few exceptions like echidnas and dolphins. In humans, they be become common by the seventh month of pregnancy, and persist throughout life even in the congenitally blind. Researchers have developed techniques to perform a full electrical sleep analysis on subjects while they are simultaneously scanned inside an MRI machine. A new study in PNAS now reports that REM sleep can be distinguished from other states of consciousness by virtue of rhythmic correlations, and anticorrelations, between different areas of the brain.
Polysomnography is a comprehensive biophysical analysis used to gauge sleep state. Most of the recorded variables, like EEG, eye movements and heart rate, are electrical in nature. In addition, many other kinds of measurements are often included like body temperature, breathing rate, or blood oxygenation. Although these variables together paint a fairly reliable picture of depth of sleep, they have little to say about what might be going on in the brain during different states of consciousness.
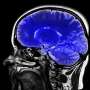
To address this problem, the researchers in the PNAS study used blood-oxygen level dependent (BOLD) MRI to assess functional connectivity between different regions of the brain. Their main finding was that the BOLD signal time series during REM sleep showed strong correlation between the thalamus and the visual cortex, and strong anticorrelation between the thalamus a region of the brain known as the posterior cingulate gyrus. Furthermore, these relations showed clear rhythmic behavior with a relatively constant period of several seconds. This temporal scale corresponds roughly to many other phasic phenomena that are seen during REM sleep.
Some of the common electrically-recorded features of REM sleep have earned names for themselves by virtue of there uniqueness. The so-called sleep spindles and k-complexes have been associated with the cessation of emg activity, and the onset of the disconnection of the brain from the musculature. At the level specific neural systems, it has long been accepted that the major monoaminergic transmitter systems of the brain take a break during REM, while the cholinergic systems become tonically active. Monoamines are those amino-acid derived transmitters that have a single amine group like noradrenaline, serotonin or histamine.
The researchers sought to partition the brain into various sensorimotor regions, and other association areas they call the default mode network (DMN). The posterior cingulate area, together with the prefrontal cortex and inferior parietal areas are said to make up this DMN. Opposite the posterior cingulate area, on the external surface of the cortex in the inferior parietal lobe, is the angular gyrus. Lying at the top of the primary fold in the brain, this area may be said to be at the convex cusp of connectivity. In other words, axons projecting from this area have more immediate short range connectivity options available to them than perhaps anywhere else in the brain. Stroke this area out, and our most fine-grained functions—mathematical, verbal and ideological—are immediately lobotomized.
As BOLD signals change relatively slowly, and can only be measured relatively slowly, they are ultimately of limited value. Uncovering the mysteries of REM sleep, and why we dream, will require much more attention to anecdote and detail. For example, it is known binocular eye movements during REM sleep can be far from conjugate in both the vertical and horizontal planes. Those creatures that show reduced levels of REM sleep have also been shown to have a smaller corpus callosum, or frequently none at all. Something about the bilateral-binocular nature of the brain seems to feature strongly in REM sleep.
At the level of dreams, it is hard to escape the idea that they have some evolved purpose, though this is not yet within the realm of fact. Many among us have dreamt of waves or waterfalls only to awake with a crushing need to visit the bathroom. Other times we teeter at the edge of a cliff, obviously standing-in for the edge of the bed, or struggle to raise a limb to defend ourself against an imaginary foe, while in reality the limb has become hypoxic under our girth. Further removed from this base physiology, our dreams may reassemble our fears and struggles, and simultaneously exaggerate and trivialize emotional events with quizzically open-ended probes.
The synchrony and interconnection of the thalamus, only accessed at low resolution in the present study, remains of central importance in the study of conscious state. Closer inspection of sensorimotor and association areas within the thalamus itself, may continue to shed more light on these issues.
More information: Rhythmic alternating patterns of brain activity distinguish rapid eye movement sleep from other states of consciousness, PNAS, Published online before print June 3, 2013, doi: 10.1073/pnas.1217691110
Abstract
Rapid eye movement (REM) sleep constitutes a distinct "third state" of consciousness, during which levels of brain activity are commensurate with wakefulness, but conscious awareness is radically transformed. To characterize the temporal and spatial features of this paradoxical state, we examined functional interactions between brain regions using fMRI resting-state connectivity methods. Supporting the view that the functional integrity of the default mode network (DMN) reflects "level of consciousness," we observed functional uncoupling of the DMN during deep sleep and recoupling during REM sleep (similar to wakefulness). However, unlike either deep sleep or wakefulness, REM was characterized by a more widespread, temporally dynamic interaction between two major brain systems: unimodal sensorimotor areas and the higher-order association cortices (including the DMN), which normally regulate their activity. During REM, these two systems become anticorrelated and fluctuate rhythmically, in reciprocally alternating multisecond epochs with a frequency ranging from 0.1 to 0.01 Hz. This unique spatiotemporal pattern suggests a model for REM sleep that may be consistent with its role in dream formation and memory consolidation.
© 2013 Medical Xpress