Epigenetic program leading to vessel differentiation
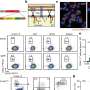
 This is an epigenetic landscape was observed in the EC generation stage. Methylation state on the histone tail on the chromatin, histone code, in endothelial cells was read via hi-speed deep sequencer. (B) Histone marking on the regulatory region of EC master transcription factors was switched following the order, (1) release the brake mark and then (2) enter the acceleration mark. (C) A new concept proposal for the EC differentiation switch. Dynamic epigenetic analysis revealed that differentiated ECs were tightly regulated via histone modification switch, which looks like a baton pass, rather than bivalent histone modifications.Adapted from: Y. Kanki, R. Nakaki, T. Shimamura, T. Matsunaga, K. Yamamizu, S. Katayama, J. Suehiro, T. Osawa, H. Aburatani, T. Kodama, et al., \"Dynamically and epigenetically coordinated gata/ets/sox transcription factor expression is indispensable for endothelial cell differentiation.,\" Nucleic acids research, Mar. 2017. DOI: 10.1093/nar/gkx159 Credit: Professor Takashi Minami")
Clarification of how human blood vessels are constructed is desperately needed to advance regenerative medicine. A collaborative research group from Kumamoto University, Kyoto University, and the University of Tokyo in Japan investigated the changes in gene functions that occur when stem cells become vascular cells. They found that the histone code, which alters the transcriptional state of the gene, changes over time as stem cells differentiate into blood vessels in response to a stimulus. Furthermore, they found that a transcription factor group essential for blood vessel differentiation (ETS/GATA/SOX) has a previously unknown role.
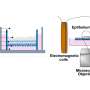
Regenerative medicine has made remarkable progress due to research with embryonic stem (ES) cells and induced pluripotent stem (iPS) cells. However, the mechanism of how blood vessels are constructed from these undifferentiated cells has not yet been clarified. During the creation of new blood vessels, the vascular endothelial growth factor (VEGF) protein differentiates stem cells into vascular endothelial cells and stimulates them to create new blood vessels. Researchers at Kumamoto University added VEGF to undifferentiated ES cells and tracked the behavior of the entire genome and epigenome changes over time in vitro.
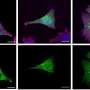
Using ES cells developed at the Center for iPS Cell Research and Application (CiRA) in Kyoto University, the research group collected RNA and histones of each cell immediately after VEGF stimulation (0 h), before differentiation (6 h), during differentiation (12 - 24 h), and after differentiation (48 h). They then comprehensively analyzed the changes in the whole genome and epigenome using next generation deep sequencing.
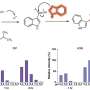
 or repressed (blue) genes by each transcription factor are shown in the box. Adapted from: Y. Kanki, R. Nakaki, T. Shimamura, T. Matsunaga, K. Yamamizu, S. Katayama, J. Suehiro, T. Osawa, H. Aburatani, T. Kodama, et al., \"Dynamically and epigenetically coordinated gata/ets/sox transcription factor expression is indispensable for endothelial cell differentiation.,\" Nucleic acids research, Mar. 2017. DOI: 10.1093/nar/gkx159")
In the process of blood vessel differentiation, the function of the protein ETS variant 2 (ETV2), which determines the differentiation into vascular endothelium, was first induced within 6 hours of differentiation stimulation. The protein GATA2, which binds to ETV2 and supports vascular endothelial differentiation, was induced immediately thereafter. Transcription factors SOX and FLI1, both important for endothelial differentiation, were induced between 12 and 24 hours. At 48 hours, after differentiation into vascular endothelium was determined, a system of transcription was established in which genes unique to vascular endothelial differentiation were induced.
Furthermore, an examination of the histone code revealed that the regulatory genomic region of the transcription factors (ETS/GATA/SOX) was found to have gradually switched from a "brake histone mark," which suppresses transcription, to an "accelerator histone mark," which activates transcription, while in the process of differentiating into the vascular endothelium. Previously, in the region that controls the function of the transcription factor that promotes differentiation from ES cells to a specific cell type, bivalent modifications of histones such as the accelerator and brake histone marks for transcription were thought to have coexisted.
In addition, when these transcription factors lose their function, terminal differentiation into the vascular endothelium (completion of differentiation) is completely suppressed, and genes that are key to differentiation into vascular endothelial cells as well as transcription factors that maintain the undifferentiated state are adversely induced. Collectively, the transcription factors (ETS/GATA/SOX) not only induce vascular endothelial differentiation, but also suppress regression to an undifferentiated state and differentiation into other ectodermal or endoderm-derived cells.
It is expected that the knowledge of the functions of these transcription factors, when combined with gene editing techniques, will allow for the efficient regeneration of blood vessels.
More information: Yasuharu Kanki et al, Dynamically and epigenetically coordinated GATA/ETS/SOX transcription factor expression is indispensable for endothelial cell differentiation, Nucleic Acids Research (2017). DOI: 10.1093/nar/gkx159