How blood vessels control their destiny

The endothelial cells that comprise blood vessels are uniquely responsive to cues from other organs, since their role is to integrate intimately into tissues and provide a means for delivery of oxygen and nutrients and waste removal. However, they also exert control over their own responses to external cues, in part by producing proteins that act to blunt the effects of the incoming signals.
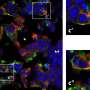
One such protein is Vascular Endothelial Growth Factor Receptor-1 (VEGFR1), which inhibits the signaling for incoming VEGF protein – a signaling protein that is crucial for new blood vessel growth but that must be tightly controlled for proper vessel growth and patterning. Acting as a "decoy," VEGFR1 dampens signaling through another receptor that delivers a positive signal for endothelial cell division and migration. Recent work in the lab of Victoria Bautch, PhD, co-director of the UNC McAllister Heart Institute, sheds light on how VEGFR1 accomplishes this. Their results were published recently in Nature Communications.
"VEGFR1 was known to function as a decoy receptor but the mechanisms underlying this function were not understood," said Bautch, who is also Beverly Long Chapin Distinguished Professor and Chair of Biology, and a member of the UNC Lineberger Comprehensive Cancer Center. "We were trying to understand what properties of VEGFR1 make it an effective decoy receptor in shaping vascular networks."
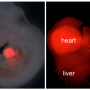
The results show that VEGFR1 is very stable, and once made can survive in endothelial cells for over a day. During this time, it slowly cycles between the outside surface and inner compartments of the endothelium, priming the endothelial cell to blunt incoming VEGF signals. If the receptor encounters VEGF while on the cell surface, VEGFR1 then undergoes degradation. The data suggest that this also degrades the VEGF protein and removes it from the system.
A process called palmitoylation makes it possible for VEGFR1 to tightly control the amplitude of incoming VEGF signal, acting as a kind of switch to regulate angiogenesis. During this process, addition of palmitic acid to the VEGFR1 protein induces stabilization and is associated with the unbound state that makes it available to serve as a decoy for VEGF. When VEGF binds to VEGFR1 it induces loss of palmitic acid and destabilization of the decoy protein.
These processes are regulated by enzymes within endothelial cells that regulate palmitoylation, and they in turn are regulated by proteins that "traffic" enzymes to the right cellular location to affect palmitoylation of the decoy VEGFR1.
"DHHC enzymes are important for palmitic acid to bind to proteins," explains Bautch. "We think a trafficking protein called RAB27 is needed to get DHHC in the same place as VEGFR1 so it can add the palmitic acid."
It is these trafficking proteins that may serve as potential therapeutic targets.
"The work we're doing is still very basic science but we are currently running experiments to test the effect loss of RAB27 has on tumor formation. If our hypothesis is correct, we predict that lack of RAB27 will mean less VEGFR1, which will mean more tumor angiogenesis and faster tumor growth."
Beyond tumor formation, these findings may also have further applications.
"Understanding more about how VEGF regulation affects disease – both excess vessel formation in tumors and perhaps a lack of vessel formation also – is the next step," said Bautch. "For example, wound healing in diabetics is compromised by lack of angiogenesis. We wonder if drugs that regulate the enzymes that add and remove palmitic acid from VEGFR1 might be an alternate and novel way to regulate blood vessel formation in humans."
More information: Joshua M. Boucher et al. Dynamic alterations in decoy VEGF receptor-1 stability regulate angiogenesis, Nature Communications (2017). DOI: 10.1038/ncomms15699