January 29, 2021 feature
How does SARS-CoV-2 evade our immune defenses?
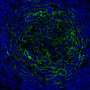
 heavily infected with SARS-CoV-2 virus particles (red), isolated from a patient sample. Image captured at the NIAID Integrated Research Facility (IRF) in Fort Detrick, Maryland. Credit: NIAID")
How does SARS-CoV-2 evade our immune defenses? It's a good question, actually. When SARS-CoV-2 defeats host defenses, we need to know how it does so, and conversely, when host defenses prevail, that also needs to be understood. The first time that a cell meets the virus, it is the local innate immune system inside the infected cell that must spring into action. By the time more diffuse bloodborne signaling and other system-wide adaptive immune responses unfold, it may be too late.
There are several kinds of cytosolic sensors in most cells that detect varieties of nucleic acids that shouldn't be there. Besides the cell's own single-stranded (ss), positive-sense (+) mRNA, the presence other cytosolic nucleic acids (like double or single strands of DNA or RNA of different senses) means that either pathogen infection, leakage of mtDNA from damaged mitochondria, or total nuclear breakdown has occurred. In any event, these are all ominous events that need to be addressed quickly before the cell is forced to roll out some kind of autodestruct plan.
The normal detector of positive sense viral RNA is the helicase RIG-1, while the helicase MDA5 senses negative sense RNA and long dsRNA. RNAs with modified nucleotides, like N-6-methyladenosine (m6A) and pseudouridine nucleotides (like those found in the Pfizer vaccine) often fail to trigger these sensors. Our own mRNA is able to escape detection through CAP-1 methylation. Occasionally, our cells make dsRNAs, and their detection is avoided by A-to-I (adenosine to inosine) modifications that are catalyzed by adenosine deaminase (ADAR1).
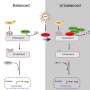
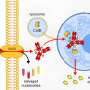
A key molecule that integrates signaling between different cytosolic nucleic acid defense molecules is STING, which stands for "stimulator of interferon genes." While its canonical function is to initiate the cGAS–STING pathway in response to cytosolic dsDNA, it is now known to participate in downstream RNA detection pathways, as well. It should not be surprising that sensing pathogenic patterns of cytosolic nucleic acids is a fundamental and conserved feature of most forms of life. In a recent paper in Trends in Biochemical Sciences, author Samuel Bakhoum pieces together the evolutionary origins of STING signaling and explores it as a therapeutic target. The fruits of this labor have broad implications for all cellular processes involving inflammation and autophagy, which extends our understanding of cell defenses beyond viral infections and into the genesis of many kinds of cancers.
The way cGAS-STING works is that upon DNA recognition, cGAS (Cyclic GMP-AMP synthase) dimerizes and stimulates formation of cyclic-GMP-AMP (cGAMP). This construct then binds and activates STING. In turn, STING phosphorylates the transcription factor IRF3 via TBK1, which then enters the nucleus to promote transcription inflammatory interferons like IFN-β. I asked Samuel why Nature has universally decided to synthesize cyclic dinucleotides as the definitive response to cytosolic nucleic acids. Specifically, whether the nucleic acids were being broken down to individual dNTPs (deoxynucleotides), converted into NTPs, and then used as the substrates for signaling cGAMP; or whether there would likely be plenty of purine NTP substrates already available in the cell. Although this question hasn't been addressed, at least to his knowledge, there may be some similarities with the common generation of cyclic nucletides in extracellular and even mitochondrial G-protein signal transduction.
To get a real feel for what STING is, it's necessary to go all the way back and try to uncover its founding purpose. An analysis of STING protein sequences has shown that its interferon signaling activities, as mediated by its C-terminal tail region, is a relatively recent addition. Its primary conserved role appears to be the generalized induction of cellular autophagy, typically in response to cytosolic dsDNA. In more primitive organisms like E. coli and V. cholera, STING's partner, cGAS, prevents the spread of bacteriophage infection to neighboring bacteria by activating phospholipase A2. This results in pore formation in the bacterial inner membrane and eventual death.
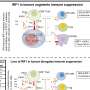
Being what it is, STING is an important target that many viruses try to inactivate in order to increase their infectivity. For example, Zika viruses, dengue virus, West Nile virus, Japanese encephalitis virus, and hepatitis C all have ways to block STING-mediated signaling. Some can cleave STING or cGAS with their proteases, and others can inhibit STING-TBK1 interaction, which would inhibit the interferon axis but spare the NF-KB arm of a response. As an enveloped, positive sense, single-stranded RNA virus, SARS-CoV-2 might draw the attention of cytoplasmic nucleic acid sensors like STING at different points in their life cycle. Its backbone is similar to our mRNA and is therefore immediately translated by the host cell. Negative-sense viral RNA is complementary to mRNA and thus must be converted to positive-sense RNA by an RNA polymerase before translation. Its dsRNA replicative intermediate form could also be a target, although the virus is regularly ensconced behind double-walled membranes.
SARS-CoV-2 itself induces many kinds of delayed cell damage, leading to release of mtDNA and other kinds of dsDNA. The SARS membrane (M) protein has been found to inhibit type I and III interferon production by targeting both RIG-1 and MDA5 through suppression of STING. Normally, activated RIG-1 engages the mitochondrial antiviral signaling protein (MAVS), whereupon the complex is modified with K63-polyubiquitin chains. These MAVS then rapidly form prion-like aggregates, which then serially convert other MAVS on the mitochondrial outer membrane. Although the full implications of this process are not yet understood, it illustrations important features of SARS-Co-V-2 that we are rapidly coming to terms with and applying in our battle against it.
More information: Nimi Vashi et al. The Evolution of STING Signaling and Its Involvement in Cancer, Trends in Biochemical Sciences (2021). DOI: 10.1016/j.tibs.2020.12.010
© 2021 Science X Network