January 25, 2021 feature
Tumor cells reveal a core syncytial drive to evade host defenses

Toxins, radiation, and normal metabolic processes together generate molecular lesions in every one of our cells to the tune of 10,000 to 1,000,000 hits per day. Fortunately, most of these code faults are immediately patched up by sophisticated DNA repair mechanisms. Any undetected damage to tumor suppressor or other key metabolic genes usually causes such a stir that the apoptotic cycle is triggered and the cell gracefully bows out of the game. If and when that fails, the cell will present antigens from any mutated proteins to the immune system so that it can be eliminated before it can go rogue.
But things do not end there. Cancer cells will keep reshuffling the deck, and eventually, they will escape or otherwise inactivate the immune cells raised against them. By the time tumors experience their first round of chemotherapy, they are already masters of deceit. The worst of the worst, when beat down, will somehow find a way to block, detoxify or pump out any drugs that challenge them. Given that tumor cells evolve from just one or a few initial clones and don't have access to the full suite of sexual or other recombination tools used by other complex, evolving systems, how do they pull off this feat of molecular ingenuity?
One answer, recently provided courtesy of Daria Miroshnychenko, et al., at the Moffitt Cancer Center in Tampa, Florida, is that tumor cells combine with each other, split the cards and then undergo their own distinct mini-evolution. This process involves a crude form of recombination and karyotype reduction until some viable, but likely heterogeneous, progeny emerge. The authors suggest that what tumors are doing is actually a form of parasexual evolution akin to that seen in other organisms like fungi, yeast or other single-celled organisms. Parasexual cycles provide for somatic variation without the overhead of full blown meiosis and dedicated germ cells.
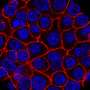
Previously, tumor cells were considered to be strictly asexual. In other words any novel genetic and epigenetic solutions they discovered would be clonal, and therefore inherited only by direct progeny. However, the frequent observations of cancer cells fusing both with normal cells, and other cancer cells, have become hard to ignore. For a locked-in cancer cell seeking greener pastures, fusion with a normal cell having a special proclivity for motility might be one way to go. The fusion process itself, especially that stimulated by certain viruses, can even be the founding tumorigenic event. In some brain tumors, like glioblastoma, it has been shown that hyperfusion to form a large, interconnected syncytium is essential for their survival against chemotherapies. It appears that this is because the network can readily redistribute mitochondria, giving select regions extra energy reserves and resistance. Other cells in the body, like muscle cells, osteoclasts, and placental cells, normally fuse into syncytia to achieve specific metabolic goals unique to their own lot in life.
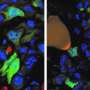
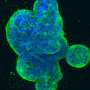
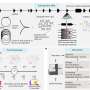
Just how common are fusion events? The researchers were able to get a handle on this question by looking at a cell line derived from a triple-negative breast adenocarcinoma. These cells lack receptors for estrogen, progesterone, and Her2 and accordingly are very invasive and difficult to treat. By labeling half of them with a green GFP construct, and half with an mCherry red construct, they could do flow cytometry with FACS fluorescent cell sorting to count cells labeled by both. Using time-lapse videography, they could also get an idea about how many doubly labeled cells actually fused, and how many were falsely positive because of incidental exchange of cytoplasm and organelles via exosomes or direct transfer from transient nanotube filaments. The researchers used an orthotopic tumor model, meaning that the cells were implanted into the normal organ of origin (orthotopically), allowing for native interaction with surrounding cells and stroma.
They also looked at fusion with other tumor lines including SUM159PT cells, and MCF-7 cells. The former have a much weaker lung colonization potential. The latter cells are more representative of many common and less aggressive tumors that do express estrogen receptors. These cells are only tumorigenic in estrogen-supplemented mice if they are engrafted into the subcutaneous fat or mammary fat pad. Without estrogen, they can be tumorigenic if engrafted intraductally.
In other recent studies, cell fusion of mesenchymal stromal cells with breast cancer cells was shown to lead to the formation at least four new and completely unique types of hybrid clones. Using experimental mammospheres, the researchers showed that each variety had their own stem character and potential for epithelial-to-mesenchymal transition. The full extent of the effects of these kinds of cell fusions in tumors remains to be seen. Other researchers have found fusion hybrid in patients who have received bone marrow transplants in the treatment of pancreatic ductal adenocarcinoma. It is known that these fusions can metastasize since melanoma cells from a transplant patient were found to have alleles from both donor and recipient.
Even more bizarre examples of exotic fusions have been documented in the recent annals of science. One woman was found to have Y chromosomes in her brain cells that apparently derived from fetal cells that managed to cross the placental barrier to enter her bloodstream while pregnant with her developing son. These cells then proved nimble enough to cross the blood-brain barrier and fuse with neurons. The fact that cells can so readily adapt themselves to these unorthodox juxtapositions of cell types and somehow knit together in an almost normal—or at least survivable—karyotype from the aftermath suggests this phenomenon may be more widespread than previously imagined.
In one of the more intriguing hypotheses for the origins of sex in the derivation of meiosis from mitosis, evolutionary biologist William Martin has suggested a multinucleated syncytial common eukaryotic ancestor. In his view, this environment was the ideal setting for the bacterially derived protomitochondrion to gain a foothold within its archaeal host. Only then would the host have sufficient energetic resources to polymerize veritable forests of tubulin filaments necessary for proper chromosome separation. The origin of eukaryotes was therefore the origin of vertical lineage inheritance, and sex was required to keep vertically evolving lineages viable by rescuing the incipient eukaryotic lineage from the pitfalls of what is commonly known as Muller's ratchet.
In light of these refined considerations, future banishment of all forms of breast cancer will likely involve taking these newfound fusion and somatic evolutionary abilities of cancer cells into account. Discrete inhibitors of cell fusion, if they are found to exist, might be one possible new avenue to investigate.
More information: Hans Zoellner et al. Potential Hydrodynamic Cytoplasmic Transfer between Mammalian Cells: Cell-Projection Pumping, Biophysical Journal (2020). DOI: 10.1016/j.bpj.2020.01.025
Sriram G. Garg et al. Mitochondria, the Cell Cycle, and the Origin of Sex via a Syncytial Eukaryote Common Ancestor, Genome Biology and Evolution (2016). DOI: 10.1093/gbe/evw136
© 2021 Science X Network